国立大学法人東海国立大学機構 名古屋大学トランスフォーマティブ生命分子研究所(WPI-ITbM)の栗原 大輔 特任講師(JSTさきがけ専任研究者)、東山 哲也 教授らの研究グループは、横浜市立大学木原生物学研究所の須崎 大地 日本学術振興会特別研究員、中部大学応用生物学部の鈴木 孝征 准教授らとの共同研究で、植物の雌の配偶子である卵細胞がつくられる様子を生きたままリアルタイムで観察(ライブイメージング)できるシステムを開発しました。
被子植物は花の中で精細胞(動物での精子)と卵細胞(動物での卵子)が出会い、受精を行います。精細胞は花粉の中でつくられますが、卵細胞は胚珠と呼ばれる将来種子となる組織の中でつくられます。しかし、胚珠はめしべの奥深くに埋め込まれているため、これまで卵細胞がつくられる様子を生きたまま観察することはできませんでした。
研究グループは、以前開発していた胚珠培養技術を用いて、胚珠の中で卵細胞がつくられる様子を生きたまま映像として捉えることに成功しました。また、卵細胞や隣接する細胞を単離して、少数の細胞で発現する遺伝子を解析する手法を確立し、卵細胞に隣接する細胞が卵細胞へと細胞運命を変化させる様子を明らかにしました。
本研究成果は、卵細胞をつくりだし受精を達成する仕組みの解明や、育種・培養技術の開発につながると期待されます。
本研究成果は、2021年3月26日付(日本時間3月27日午前3時)米国科学雑誌『PLOS Biology』電子版に掲載されました。
【ポイント】
- 植物の卵細胞がつくられる様子を生きたままリアルタイムで観察することに成功した。
- 顕微鏡下で配偶子や助細胞を取り分けて、それぞれの遺伝子発現情報を解析する実験系を確立し、卵細胞に隣接する細胞が卵細胞のように変化していく様子を明らかにした。
- 卵細胞を維持し受精を達成する仕組みの解明、育種・培養技術の開発への貢献が期待される。
【研究背景】
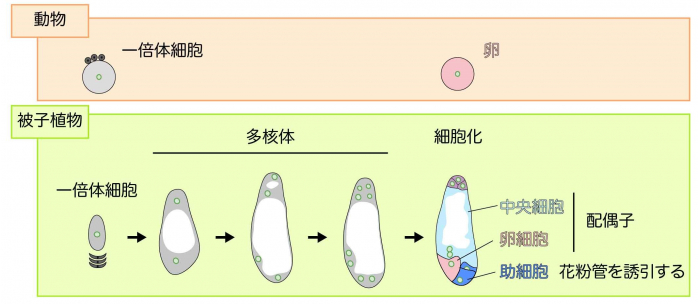
有性生殖を行う生物は、雄と雌の配偶子注1)が受精することによって、子孫を残します。雌の配偶子の場合、動物では、減数分裂によって生じた一倍体細胞が直接卵子(雌性配偶子)になりますが、多くの被子植物では、減数分裂によって生じた一倍体細胞(配偶体注2))は、さらに3回の核分裂を進行させ、多核体注3)の状態で発生を続けます。その後、細胞分裂し、2つの配偶子である卵細胞・中央細胞、また花粉管を誘引する助細胞といった、それぞれ異なる役割を担う細胞をつくり出します(図1)。しかし、多核体細胞から7つの細胞へと分裂するときに、どのように精確に2つの雌性配偶子とそれ以外の細胞をつくり出しているのか、その詳細はまだ明らかになっていません。細胞内の核の位置がそれぞれの細胞運命に重要だと考えられていますが、配偶子形成は花の奥深くにある胚珠の中で起こっているため、これまで一連の過程をリアルタイムで観察することが困難でした。研究グループは、これまでに配偶体の一部がむき出しになったトレニアという植物を用いてその発生過程の一部の観察に成功していましたが、全過程や細胞運命が決まる様子は不明でした。
減数分裂後の一倍体細胞は、動物では直接卵子となりますが、多くの被子植物では3回の核分裂を進行させ、多核体の状態で発生を続けます。8核になった後、細胞膜を形成することで(細胞化)、2つの配偶子である卵細胞・中央細胞、また花粉管を誘引する助細胞をつくり出します。
【研究内容】
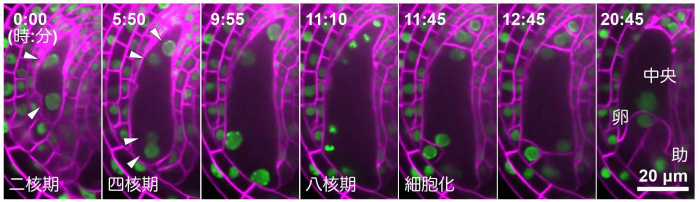
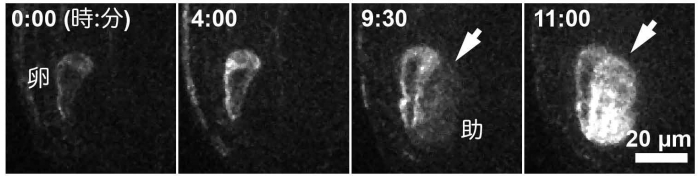
研究グループは、これまでに、モデル植物であるシロイヌナズナを用いて、将来種子となる母体組織(胚珠注4))をめしべから取り出して、胚珠の中で受精卵の分裂と、それに続く胚発生を再現することに成功していました。今回、この胚珠培養技術を用いて、シロイヌナズナの雌性配偶子がつくられる様子を生きたままリアルタイムで観察することに成功しました。観察の結果、減数分裂によって生じた核は分裂して、細胞の両極へと離れていきました。さらに分裂した4つの核は、細胞の周縁部に沿って一列に並びました。そして8つの核に分裂後、細胞膜が形成し、それぞれ2つの配偶子(卵細胞と中央細胞)と付随する細胞がつくられていく様子が詳細に明らかとなりました(図2)。157例、リアルタイムで観察したところ、配偶体細胞の花粉管が侵入する側に近い核から、必ず助細胞・卵細胞・中央細胞となり、細胞内の核の位置とその後の細胞運命に非常に高い相関があることが改めて示されました。
緑色は細胞核を、ピンク色は細胞膜を、矢じりは配偶体細胞の核を示しています。配偶体細胞の中に核が2つある二核期から観察を始めており、四核期・八核期と核分裂を繰り返した後、細胞膜を形成し細胞化することで、配偶子である卵細胞・中央細胞、隣接する助細胞をつくります。数字は観察開始からの時間を表しています。
続いて、それぞれの細胞運命注5)がいつ決定されるのかを明らかにするために、助細胞の分化と機能に重要な特異的転写因子であるMYB98の発現開始時期を解析したところ、8核に分裂し細胞膜を形成した後、早い時期からMYB98の発現が助細胞で検出されました。卵細胞特異的遺伝子も同様に早い時期から卵細胞で検出されたため、配偶子や助細胞の細胞運命は、細胞膜形成後ごく初期、あるいはそれよりも前に決まっている可能性が考えられます。
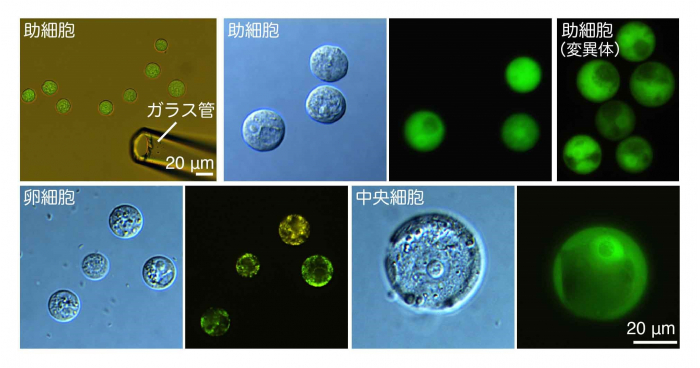
さらに研究グループは、myb98変異体において、助細胞内の核動態が不安定になり、細胞が肥大することを見いだしました。この核動態の異常が細胞運命にどのように影響しているのかを明らかにするために、配偶子や助細胞を単離し、遺伝子発現解析を行いました。配偶子や助細胞は胚珠の奥に埋め込まれているため、細胞特異的な遺伝子発現を調べるには、それぞれの細胞を綺麗に取り出して、限られた少数の細胞から発現する遺伝子の情報を得なければなりません。そこで、研究グループは顕微鏡下で配偶子や助細胞を取り分けて、それぞれの遺伝子発現情報を解析する実験系を確立することで、各細胞の特異的遺伝子を明らかにしました(図3)。
胚珠を細胞壁分解酵素で処理することで配偶子と助細胞を単離する手法を確立しました。これにより蛍光タンパク質で標識した各細胞を顕微鏡下で集めて遺伝子発現を調べることに成功しました。
myb98変異体の助細胞の遺伝子発現を野生型の助細胞と比べたところ、驚くべきことに、変異体の助細胞では多くの卵細胞特異的な遺伝子が高発現することが分かりました。そのうちのひとつである卵細胞特異的タンパク質SBT4.13の発現をmyb98変異体でリアルタイムに観察したところ、細胞膜形成後、卵細胞でSBT4.13が検出された後、比較的早い時期に助細胞でSBT4.13の異所的発現が見られました(図4)。
図4 変異体助細胞における卵細胞特異的遺伝子の異所的発現
myb98変異体で卵細胞特異的タンパク質SBT4.13の発現を解析したところ、細胞化後、卵細胞で発現が検出されましたが、その後、助細胞で異所的発現が検出されました。数字は観察開始からの時間を表しています。
研究グループは、以前、トレニアという植物を用いて、卵細胞を破壊したときに、助細胞が卵細胞様の挙動を示すことを発見しています。今回の観察結果から、シロイヌナズナにおいて、助細胞へ分化できない細胞で卵細胞特異的遺伝子が発現していたことから、植物において、配偶体細胞運命の初期設定は配偶子であり、助細胞ではその運命が抑えられていること、配偶子に異常が見られたときには助細胞で抑えられていた配偶子の運命が発揮されると考えられます。
【成果の意義】
配偶子のような重要な細胞を維持するといったこれらの結果は、移動することのできない植物がさまざまな環境変化に対応するために柔軟に細胞運命を変化させる、驚くべき生存戦略の一端を示した研究結果と言えます。今後は、どのように細胞の運命転換を誘導しているのか、分子メカニズムを明らかにしていくことが焦点になります。細胞の運命転換を可能にする分子メカニズムを明らかにすることで、受精効率の向上など、植物育種技術の開発への貢献も期待されます。
また、配偶子や助細胞の細胞運命はかなり早い段階で決まっていますが、多核体細胞内の核の位置が細胞運命にどのように影響しているのかは依然として不明です。本研究成果により確立した、植物雌性配偶体のリアルタイム観察、細胞単離、少数細胞の遺伝子発現解析といった技術を駆使することで、世界中で植物配偶子研究が加速すると期待されます。
【用語説明】
注1)配偶子
受精して次世代を産み出す生殖細胞のこと。植物の精細胞と卵細胞、動物では精子と卵子に該当する。
注2)配偶体
植物において配偶子をつくり生殖を行う世代の生物体。一倍体世代であり、雌の配偶体は胚のうとも呼ばれる。
注3)多核体
1つの細胞中に多数の核が存在する状態。一般的に細胞は1つの核をもつが、多核体となる細胞では、核が分裂しても細胞質は分裂しない細胞分裂を繰り返すことで、複数の核をもつ細胞になる。植物では初期の胚乳が多核体を形成する。
注4)胚珠
めしべの奥深くに位置する種子の元となる組織。胚珠の中にある卵細胞と花粉管によって運ばれてきた精細胞が受精することによって、受精卵がつくられる。その後、受精卵が分裂を繰り返し胚発生が進行する。
注5)細胞運命
ある細胞が将来どのような細胞種へ分化するのかという運命のこと。
【論文情報】
雑誌名:PLOS Biology
論文タイトル:Dynamics of the cell fate specifications during female gametophyte development in Arabidopsis (シロイヌナズナ雌性配偶体形成における細胞運命決定のダイナミクス)
著者:Daichi Susaki, Takamasa Suzuki, Daisuke Maruyama, Minako Ueda, Tetsuya Higashiyama, Daisuke Kurihara
(須崎 大地、鈴木 孝征、丸山 大輔、植田 美那子、東山 哲也、栗原 大輔)
DOI:10.1371/journal.pbio.3001123
【著者所属】
名古屋大学トランスフォーマティブ生命分子研究所 東山 哲也、栗原 大輔
JSTさきがけ 栗原 大輔
名古屋大学大学院理学研究科 植田 美那子、東山 哲也
東京大学大学院理学系研究科 東山 哲也
横浜市立大学木原生物学研究所 須崎 大地、丸山 大輔
中部大学応用生物学部 鈴木 孝征