~助細胞による誘引ペプチド分泌のメカニズムを解明~
横浜市立大学 木原生物学研究所 須崎大地特任助教、泉理恵さん(当時大学院生)、丸山大輔准教授らと、名古屋大学 大井崇生助教、武内秀憲特任助教、ケンタッキー大学 河島友和准教授、東京大学 東山哲也教授らの共同研究グループは、モデル植物のシロイヌナズナを用いて、胚珠
*1内にある助細胞
*2(メス)のアクチン繊維
*3が、花粉管(オス)を導くための花粉管誘引ペプチドの分泌を制御することを明らかにしました(図1)。
雌しべの中の胚珠には、雌雄の相互作用に重要な花粉管誘引ペプチドを分泌する2つの助細胞があります。助細胞は繊形装置とよばれる細胞膜と細胞壁が複雑に陥入した特殊な構造から花粉管を誘引する小さなタンパク質(ペプチド)を分泌しますが、その分泌の仕組みは明らかでありませんでした。本研究では遺伝学的、薬理学的手法によって、助細胞のアクチン繊維が誘引ペプチドの極性分泌を担っていることを解明しました。さらに、花粉管の放出で一時的に消失したアクチン繊維が、経時的に回復することを発見しました。これは受精失敗時に残った助細胞が誘引を再開するために働くと考えられます。
本研究成果は、米国の科学雑誌「The Plant Cell」オンラインアドバンス版に掲載されました。(2022年12月23日)
研究成果のポイント
- 顕微鏡観察によって助細胞の繊形装置の形態と細胞骨格の配向を明らかにした
- 助細胞のアクチン繊維が花粉管誘引ペプチドの極性分泌を制御する
- 花粉管の放出で消失した助細胞のアクチン繊維が再生して花粉管誘引を再開する
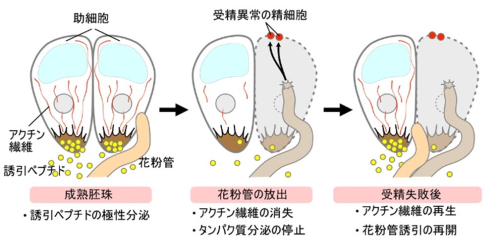
図1 助細胞のアクチン繊維による花粉管誘引ペプチド分泌の制御
左:成熟胚珠の助細胞のアクチン繊維と誘引ぺプチドの分泌。
中:花粉管の放出時に助細胞ではアクチン繊維が消失してタンパク質の分泌が停止する。
右:受精失敗後にアクチン繊維が再生して花粉管誘引が再開する。
研究背景
穀類など被子植物の種子の形成には、2つの精細胞が卵細胞と中央細胞とそれぞれ受精する重複受精が不可欠です。雌しべの胚珠の中にある2つの助細胞は花粉管誘引ペプチドを分泌することにより、花粉管を胚珠ま9で誘導する等の雌雄の相互作用に重要な役割を果たします。助細胞の先端には複雑な細胞膜の陥入と厚い細胞壁で構成される繊形装置という構造があり、核はこの近く偏って配置します。誘引ペプチドなどは繊形装置経由して分泌されると考えられています。これまでの形態学的な研究では、繊形装置から放射状に広がる微小管*4と、助細胞の縦方向に分布するアクチン繊維が観察されていました。こういった助細胞の特徴的な形態と細胞骨格の配向は、細胞機能に重要であると考えられてきましたが詳細は不明でした。
研究内容
丸山准教授の研究グループでは、助細胞の形態と機能のつながりを明らかにするために、電子顕微鏡で繊形装置の形態観察を、光学顕微鏡で微小管またはアクチンの経時的な動態変化を観察しました。また、微小管またはアクチンを遺伝学的、薬理学的な方法で重合阻害した際の細胞の形態と花粉管の誘引を調べました。微小管の破壊では、繊形装置の細胞膜の陥入が少なくなりましたが、花粉管の誘引に異常は見られませんでした。一方、アクチンを破壊すると繊形装置の形態異常だけでなく、花粉管誘引ペプチドの分泌が阻害されて発生異常を示す種子が見られました(図2)。受精異常を示す精細胞膜タンパク質の変異体の花粉管を受容して残った一方の助細胞では、一時的なアクチン繊維の消失が観察されました。さらに、経時的な観察によって再びアクチン繊維が観察されました(図3)。野生型においても受精後に残った助細胞でアクチン繊維が観察された。このアクチン繊維の再形成は、受精失敗時に再び花粉管を誘引するために機能すると考えられます。
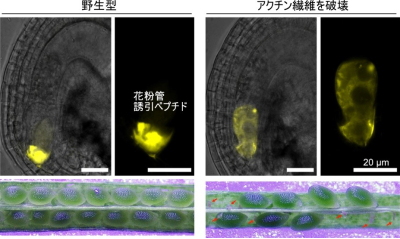 図2 助細胞のアクチン繊維を破壊すると花粉管誘引ペプチドの局在が変化し種子の発生異常が見られた
図2 助細胞のアクチン繊維を破壊すると花粉管誘引ペプチドの局在が変化し種子の発生異常が見られた
左:野生型の助細胞では花粉管誘引ペプチドが繊形装置に局在する
右:アクチン繊維を破壊すると花粉管誘引ペプチドが繊形装置に局在できなくなり、受精できず種子が正常に形成されなくなる(赤矢印)
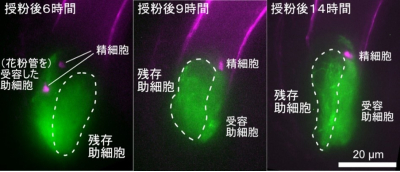
図3 受精失敗後に残った助細胞では消失したアクチン繊維が回復する
緑色:アクチン、マゼンタ:精細胞
左:授粉後6時間では残存助細胞ではアクチン繊維が消失する
中:授粉後9時間ではほとんどアクチン繊維は見られない
右:授粉後14時間ではアクチン繊維の回復が見られた
今後の展開
本研究によって重複受精の重要なステップである花粉管誘引が助細胞のアクチン繊維によって制御されていることが明らかになりました。今後は、極性分泌や細胞形態の制御遺伝子の同定など助細胞の基礎的な知見の蓄積を通じて、胚珠に同時に複数の花粉管が誘引されないようにする「多花粉管拒否機構」が明らかになると期待されます。これらの知見を基に受精の過程を操作することで、稔実率の向上や育種技術の開発につながることが期待されます。
研究費
本研究は、科学研究費助成事業(JP17H05846, JP19H04869, JP20H03280, JP20H05778, JP20H05781, JP19K16172, JP22K15145, JP18K14729, JP20K15817, JP22H05172, JP22H05175)、豊秋奨学会研究費助成事業、米国国立科学財団助成金(IOS-1928836)の支援を受けて実施されました。本研究は文部科学省ナノテクノロジープラットフォーム課題として名古屋大学微細構造解析プラットフォームの支援を受けました。
論文情報
タイトル:F-actin regulates polarized secretion of pollen tube attractants in Arabidopsis synergid cells
(シロイヌナズナの助細胞ではアクチン繊維が花粉管誘引物質の極性分泌を制御する)
著者:須崎 大地、泉 理恵、大井 崇生、武内 秀憲、Ji Min Shin、杉 直也、木下 哲、東山 哲也、河島 友和、丸山 大輔
掲載雑誌: The Plant Cell
DOI:
https://doi.org/10.1093/plcell/koac371
用語説明
*1 胚珠:被子植物の雌しべの中にある組織で、受精後に種子になる。卵細胞や中央細胞という配偶子を含む多細胞で構成される。
*2助細胞:胚珠内で卵細胞の両隣に位置する2つの細胞で、花粉管を誘引するペプチドを分泌するなど雌雄の相互作用に重要な役割を果たす。
*3アクチン繊維:細胞骨格の一種で、細胞の形の維持などにはたらく。アクチンタンパク質が重合してできた繊維状の構造で細胞内ネットワークを作っている。
*4微小管:チューブリンタンパク質が重合しできた管状構造の細胞骨格。細胞の形態形成や細胞分裂時の染色体分離に重要な役割を果たす。