2024年8月20日
甲南大学
東京農業大学
動物と植物に共通した高温耐性と低温耐性の仕組みを発見
(高温耐性を強化し低温耐性を抑える遺伝子を発見)
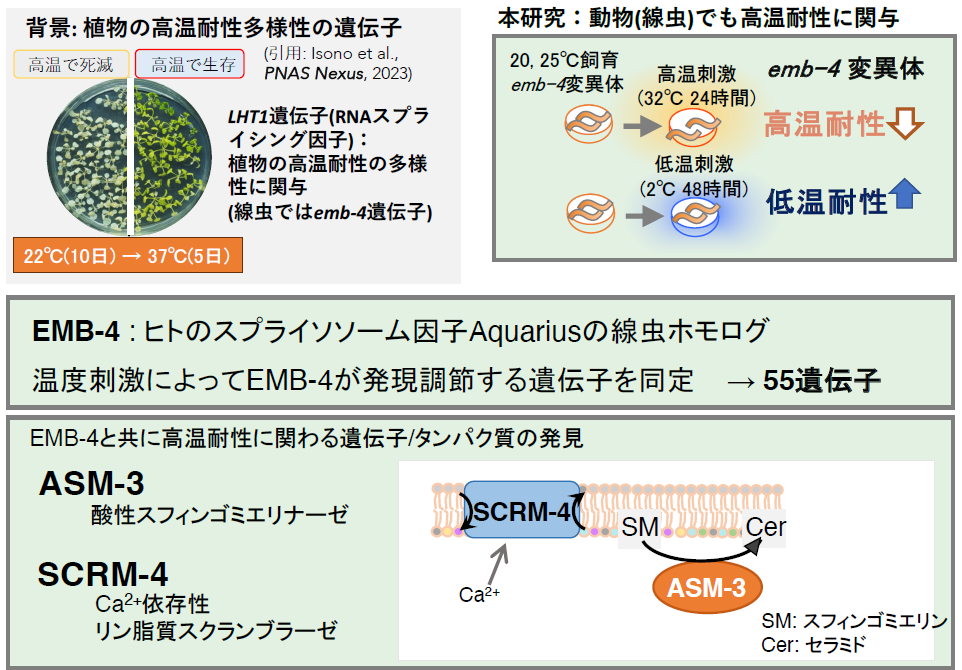
甲南大学大学院自然科学研究科の久原篤 教授、太田茜 特任研究准教授、佐藤夕希 修士課程院生(研究当時)、東京農業大学 太治輝昭 教授、礒野一帆 博士後期課程院生(研究当時)らの研究チームは、「動物と植物に共通した高温耐性と低温耐性の仕組み」を線虫とシロイヌナズナの解析から明らかにしました。
これまでに久原篤 教授と太田茜 特任研究准教授らの研究チームはシンプルな実験動物である線虫C. エレガンス注1)の温度耐性を解析してきました。また、太治輝昭 教授らの研究チームは、世界各地から単離したシロイヌナズナの高温耐性の多様性に関わる遺伝子を解析してきました。今回、昨年発見されたシロイヌナズナの高温耐性を増強するLHT1遺伝子注2)が、線虫ではemb-4という遺伝子であること、emb-4遺伝子の機能が低下した線虫の変異体では高温耐性が弱くなり、逆に低温耐性が強くなることを発見しました。emb-4遺伝子から生成されるEMB-4タンパク質は、ヒトではAQR注3)と呼ばれ多くの遺伝子のイントロン注4)に結合して、それら遺伝子産物が正常に生成されるためのスプライシング反応注5)を調整しています。EMB-4が調節する遺伝子をRNAシーケンシング解析注6)を用いて同定したところ、脂肪酸代謝に関わる酸性スフィンゴミエリナーゼ注7)や生体膜の局所的な構造に関わるリン脂質スクランブラーゼ注8)が見つかりました。それらの変異体線虫では高温耐性が強くなりました。
以上より、動物(線虫)と植物(シロイヌナズナ)という全く質的に異なる生物種に共通した高温耐性に関わる遺伝子が明らかになり、線虫では脂肪酸の代謝や生体膜の構造に関わる遺伝子を調節していました。emb-4遺伝子の類似遺伝子は、ヒトを含む動物から植物まで広く存在しているため、解析を進めることで、地球温暖化による食糧危機に貢献できる高温に強い家畜や農作物の開発に繋がることが期待されます。本研究の成果は、2024年8月8日に、英国科学雑誌「PNAS nexus」のオンライン版に掲載されました。
|
<研究の背景と経緯>
『温度情報は生物が生存していく中で重要な環境因子です。例えば、人間では体温がわずか3℃低下し33℃になると仮死状態になることや、病気の発生率や死亡率は気温変化が大きい時季に増加します。温度が関わる病気は多数あり、急激な温度変化によるヒートショック死や低体温症が社会問題となっており、これらにより亡くなる方の数は交通事故による死亡数よりも多いという報告があります。地球規模でも、温暖化や局所的な寒冷化により生物種の3割が絶滅の危機に瀕すると報告されており、生物の温度適応は世界的な課題となっています。
生物は生育環境から絶え間なく温度情報を受け取り、それに適応することで生存してきました。そのため、生物は温度変化を感知して、それに対応する生体メカニズムを持っています。例えば人間では自律的な体温調節機構が存在し、深部体温と環境温度の変化をそれぞれ感知して情報を統合し、適切な体温調節を行います。線虫からヒトまで多くの動物で進化的に保存された温度センサー分子として、TRP(Transient receptor potential)イオンチャネルが知られており、様々な細胞や組織で発現し、細胞内カルシウム濃度を増加させることにより多くの熱感覚応答に関与しています。
生物の温度応答メカニズムの解明は多方面から望まれている一方で、ヒトの体は約37兆個の細胞で構成されているため、その細胞ネットワークは非常に複雑です。そこで本研究チームは、動物の温度応答メカニズムの解明に向けて、ヒトと相同な遺伝子を多く持つシンプルなモデル実験動物である線虫
C. エレガンスを用いました。
C. エレガンスは959個の細胞を持ち、そのうち302個が神経細胞です。
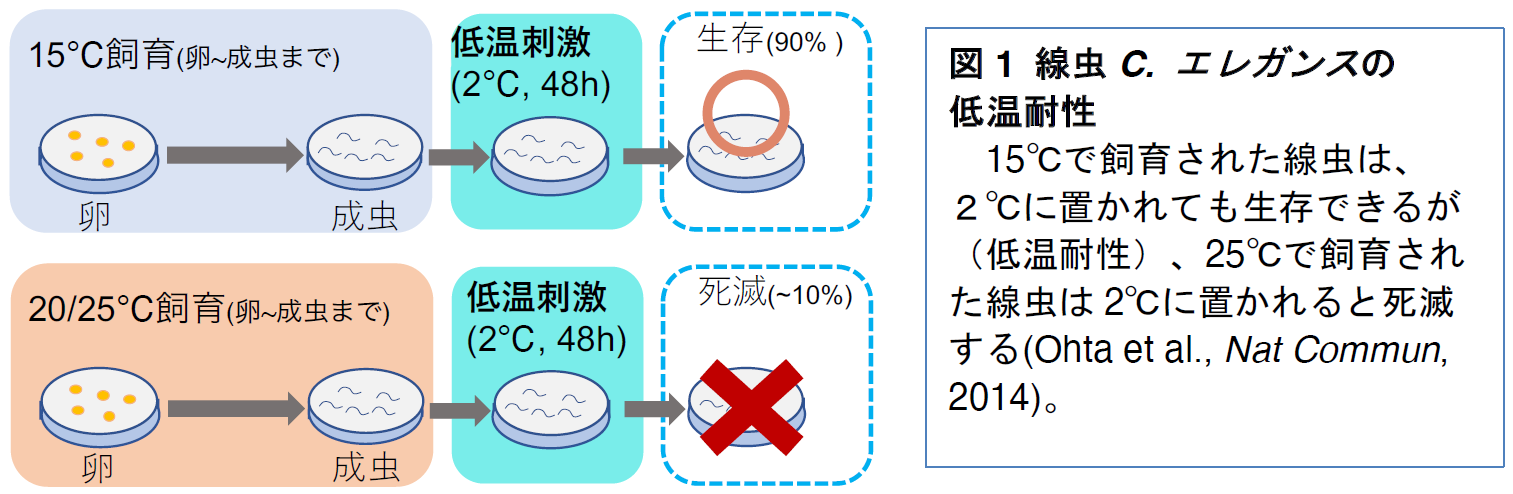
C. エレガンスの温度への適応機構として、これまでに温度順化現象が確認されています。温度順化現象とは、飼育環境の変化に合わせて低温(2℃)への耐性が変化する現象を指します
(図1)。具体的には、15℃で飼育した線虫は低温耐性を持ち、2℃に移されても生存可能ですが、25℃で飼育した線虫は低温耐性を持たず、2℃では死滅します
(図1)。
この低温耐性を指標に、温度応答に関わる分子や神経細胞の解析が進められてきました。』
(「研究の背景と経緯」の文章の『』で囲われた部分は、過去のプレスリリース「温度への慣れに関わる脳・腸連関 令和4年7月27日 甲南大学」から引用(一部改変))
これまでに、低温耐性や温度順化に関わる温度受容ニューロンとして、頭部に位置する3種類の温度受容ニューロンが見つかっています。温度受容ニューロンの一つが温度を受容すると、シナプスからインスリンを分泌して腸で受け取り、腸内の不飽和脂肪酸合成を抑制することで低温耐性を負に制御します。また、低温時には腸内の中性脂肪の量を増やします。一方、高温耐性に関しては、熱ショックタンパク質などが必要なことが知られています。しかし、低温耐性と高温耐性に共通した遺伝子は十分には分かっていませんでした。
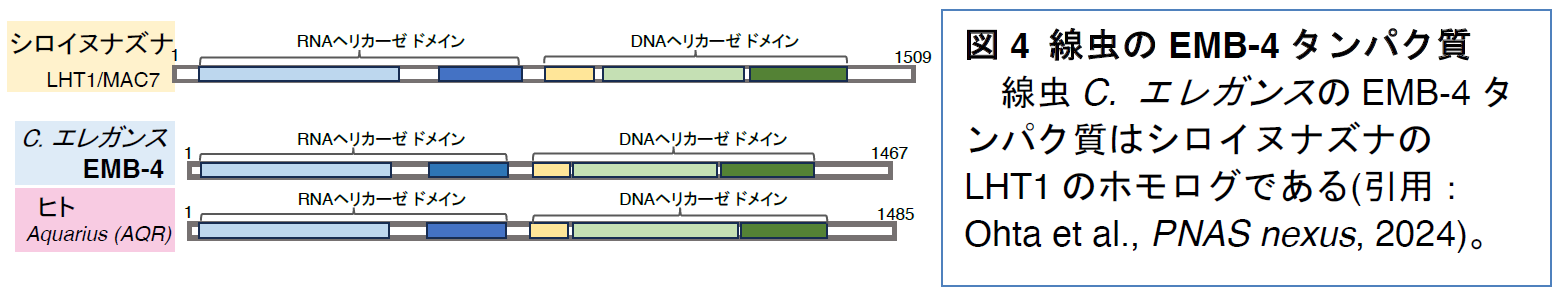
高温耐性は、植物においても解析が進んでいます。そのなかでも、モデル植物であるシロイヌナズナは、世界各地に2000種類以上の野生系統が存在します。そのうち高温耐性の低い系統と高い系統の責任遺伝子として、Long-term Heat Tolerance1 (LHT1) /MAC7を同定しました
(図2)。LHT1/MAC7遺伝子は、mRNAのスプライシングに関与するRNAヘリカーゼ
注9)をコードし、LHT1/MAC7はヒトのAquarius (AQR)の相同遺伝子でした。一方で、これらの遺伝子がヒトを含む動物の高温耐性や低温耐性に関与しているかは分かっていませんでした。
<研究の内容>
線虫C. エレガンスの野生株N2は飼育温度依存的な高温耐性を示す
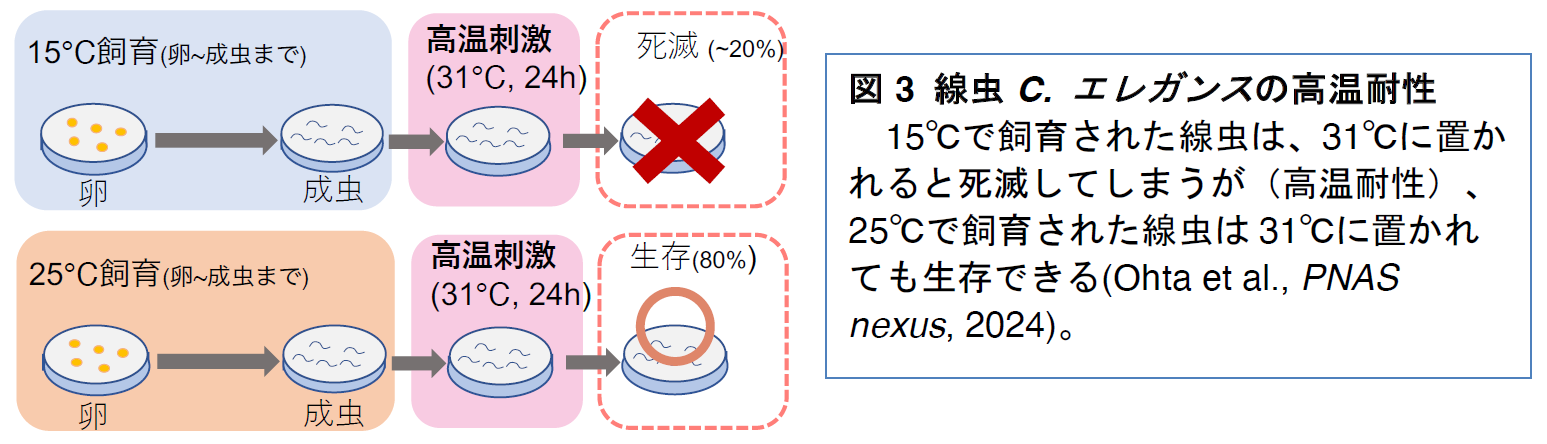
線虫
C. エレガンスの野生株N2が、過去の飼育温度に依存して高温耐性を変化させるか調べたところ、15℃で飼育された個体の大部分は31℃で死亡し、25℃で飼育された個体は31℃でも生存し、20℃で飼育された個体は半分ほどが生存できる表現型を示しました
(図3)。これらの結果は、高温にさらされた後では、より高い温度に曝されても生存できるようになることを示しており、高温耐性の獲得や喪失が過去の飼育温度に依存することを示唆しています。
植物の高温耐性に関わる遺伝子が線虫の高温耐性と低温耐性にも関わる
近年見つかったシロイヌナズナの高温耐性の多様性に関わるLHT1遺伝子が動物の温度耐性にも関わるか線虫を用いて解析しました。LHT1タンパク質はRNAヘリカーゼドメイン注9)とDNAヘリカーゼドメインをもつ遺伝子のスプライシングに関わるスプライソソーム因子注10)で、ヒトのAQRのホモログです。C. エレガンスではemb-4遺伝子によってコードされているEMB-4タンパク質のホモログであることが分かりました(図4)。
そこで、
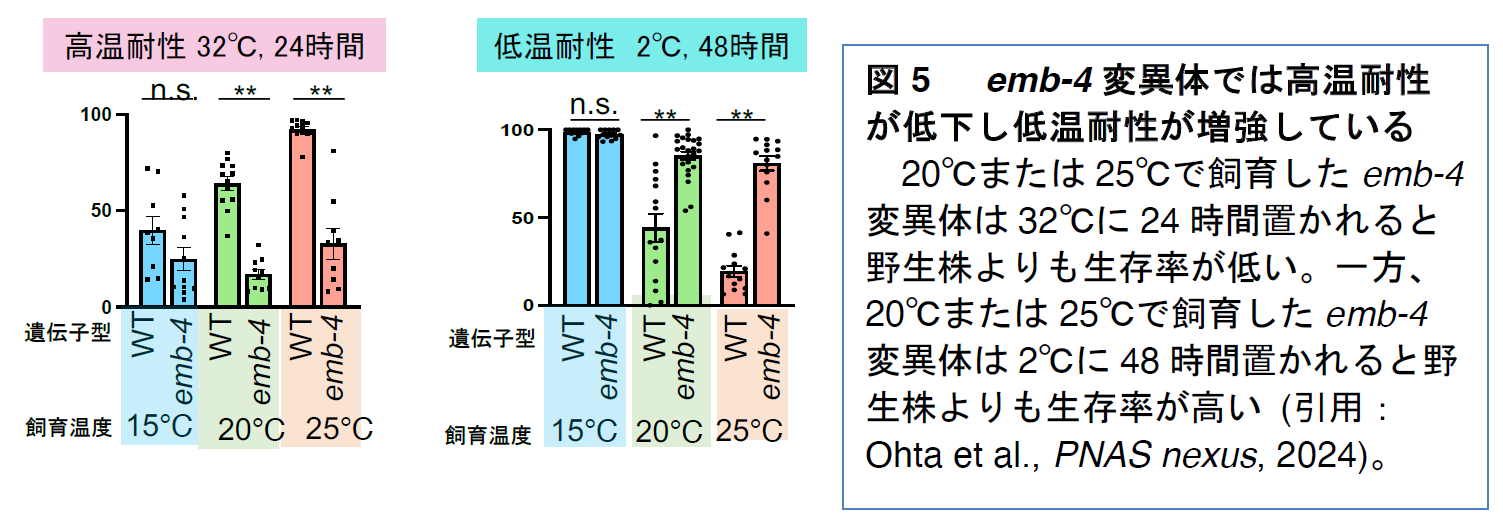
emb-4変異体の高温耐性を測定したところ、20℃または25℃で飼育した後に32℃に24時間置かれた際の生存率が野生株よりも低下していました
(図5)。つまり、EMB-4は線虫の高温耐性を正に調節していると考えられます。一方、
emb-4変異体の低温耐性を測定したところ、20℃または25℃で飼育した後に2℃に48時間置かれた際の生存率が野生株よりも上昇していました
(図5)。つまり、EMB-4は線虫の低温耐性を負に調節していると考えられます。これらのことから、野生株のEMB-4タンパク質は高温耐性を促進させ、低温耐性を抑制させる機能があると考えられます。
高温刺激や低温刺激を与えた際にemb-4変異体において発現変動する遺伝子を同定
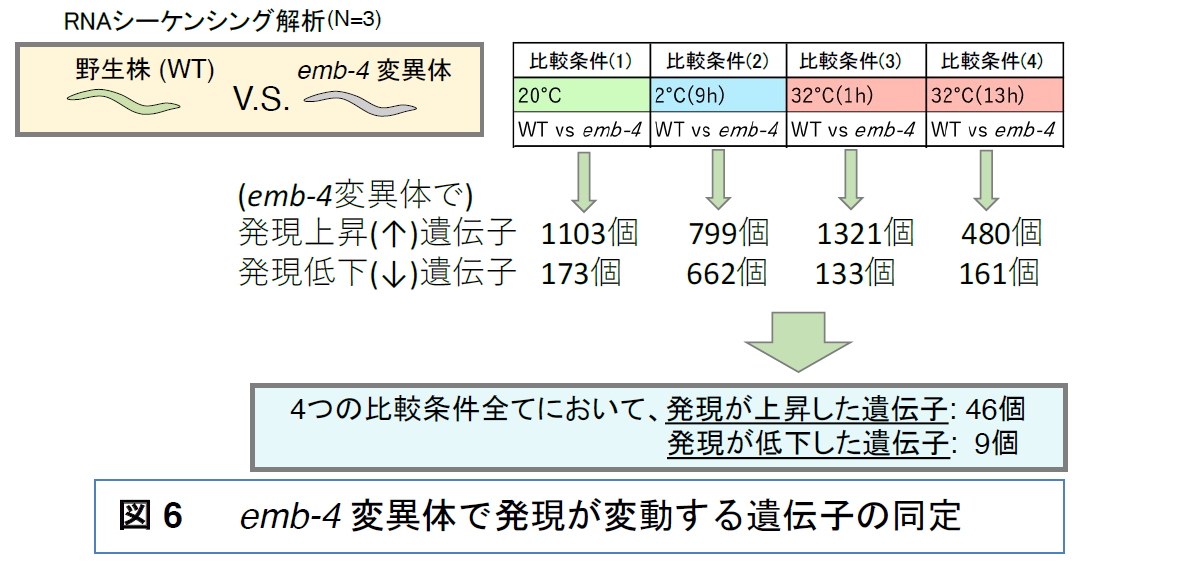
高温耐性と低温耐性においてEMB-4タンパク質がどのような遺伝子の発現に影響を与えるかをトランスクリプトーム解析から調べました。下記の(1)〜(4)の4つの比較条件で解析をし、それぞれ発現が上昇と低下している遺伝子を見つけました
(図6)。
(1) 20℃で飼育し続け、温度変化を与えなかった
emb-4変異体と野生型株で発現レベルが異なった遺伝子[20℃ =発現上昇(↑):1103個の遺伝子、発現低下(↓):173個の遺伝子]
(2) 20℃飼育後に2℃に9時間置いた場合に、
emb-4変異体と野生型株で発現レベルが異なった遺伝子[2℃(9h) =↑:799個、↓:662個]
(3) 20℃飼育後に32℃に1時間置いた場合に、
emb-4変異体と野生型株で発現レベルが異なった遺伝子[32℃(1h) =↑:1321個、↓:133個]
(4) 20℃飼育後に32℃に13時間置いた場合に、
emb-4変異体と野生型株で発現レベルが異なった遺伝子[32℃(13h) =↑:480個、↓:161個]
上記の4つの条件で
共通して発現が変動する55個の遺伝子を見つけた[共通 =↑:46個、↓:9個]
(図6)。
高温耐性に関わる新しい遺伝子を同定
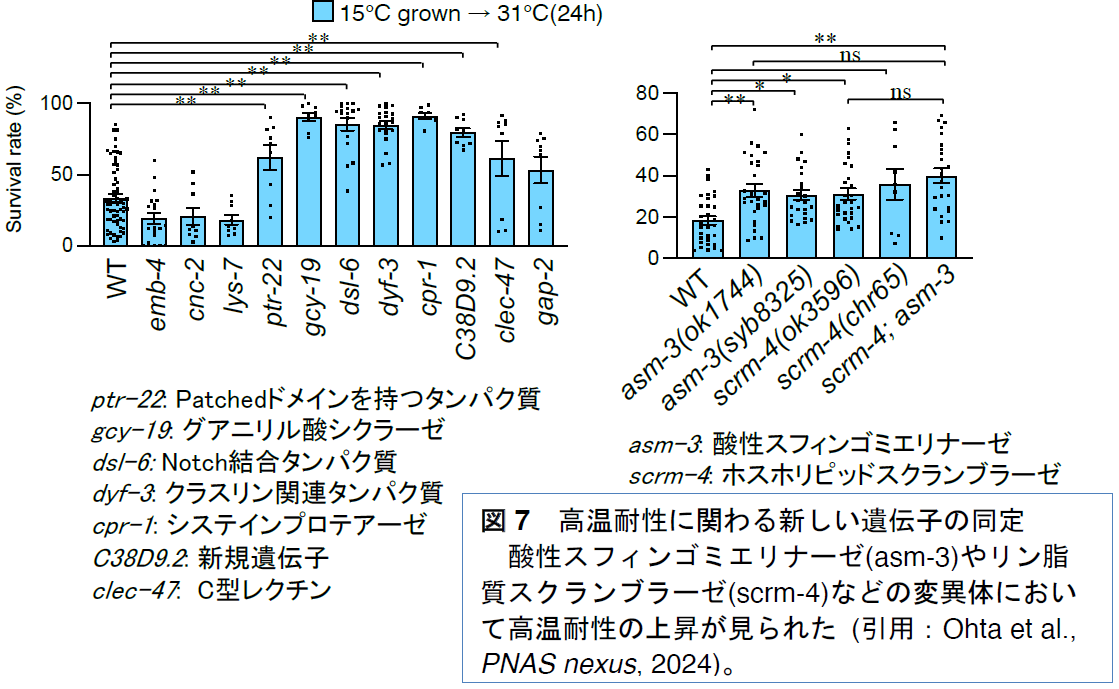
emb-4変異体において発現が変動していた55個の遺伝子のうち変異体の存在した12個の遺伝子について、高温耐性と低温耐性への関与を調べました
(図7)。その結果、脂肪酸代謝に関わる酸性スフィンゴミエリナーゼ(
asm-3)や膜リン脂質の局所的な構造に関わるリン脂質スクランブラーゼ(
scrm-4)の変異体において高温耐性の増強が見られました。さらに、神経系で機能するグアニリル酸シクラーゼ(
gcy-19)やクラスリン様タンパク質(
dyf-3)の変異体などでも高温耐性の増強が見られました。また、機能が未知の新規の遺伝子(
C38D9.2)の変異体においても高温耐性が増強していました。これらのうち、
dyf-3と
C38D9.2の変異体では低温耐性も増強していました。
酸性スフィンゴミエリナーゼ (ASM) は、細胞膜やリソソームなどの脂質二重膜に存在し、スフィンゴミエリンを加水分解してセラミドとリン酸化コリンを生成します。ASMによって生成されるセラミドを含むスフィンゴ脂質は、細胞膜シグナル伝達などの中心となる脂質ラフト(lipid raft)の形成に関与します。また、
asm-3変異体では2種類のオメガ-3長鎖脂肪酸が過剰に蓄積することが報告されています。以上のことから、ASM-3を介した細胞膜シグナル伝達や脂肪酸代謝が高温耐性に重要である可能性があります。
リン脂質スクランブラーゼ (SCRM) も、細胞膜やミトコンドリアなどの脂質二重膜に存在し、リン脂質(ホスファチジルセリン)を内膜から外膜へと移動させます。SCRMの主な活性化因子はCa2+であり、これは血液凝固、アポトーシス、マイトファジーシグナルによって導入されます。最近の報告では、線虫においてSCRMであるATG-9が脂肪滴からの脂質動員に関与していることが示唆されています。これらのことから、Ca2+によって調節される膜リン脂質の局所的な構成が、SCRM-4を介した高温耐性の制御に関与している可能性があります。
なお、これまでにEMB-4は胚発生に関わることが知られていましたが、今回、EMB-4が神経系の遺伝子の発現にも影響を与えるという新たな機能も見つかりました。
<今後の展開>
本研究から、これまでに植物の高温耐性に関わることが報告されたスプライソソーム因子が、動物である線虫の高温耐性と低温耐性にも関わることが見つかりました。さらに線虫における該当遺伝子(
emb-4)が、脂肪酸代謝に関わる酸性スフィンゴミエリナーゼや膜リン脂質の局所的な構造に関わるリン脂質スクランブラーゼなどの遺伝子発現に影響を与えることで、高温耐性を制御していることが分かりました。
今回、動物と植物の高温耐性を制御する共通の遺伝子が見つかってきました。線虫の
emb-4遺伝子の類似遺伝子は、ヒトを含む動物から植物まで広く存在しているため、今後、さらに解析を進めることで、高温に強い家畜や農作物の開発に繋がることが期待されます。
<研究支援>
本研究は、日本学術振興会科学研究費助成事業(24H01255、24H02022、24K02070、21K06275、24K09538、23H00334、23H04206)、甲南学園平生太郎基金科学研究奨励助成、日本医療研究開発機構(AMED)革新的先端研究開発支援事業(PRIME)(24gm6510004h0004)、内藤記念科学振興財団、武田科学振興財団、旭硝子財団、光科学技術研究振興財団、山田科学振興財団、東レ科学振興会、ノバルティス科学振興財団の研究費によって行われました。
<用語解説>
注1)
C.エレガンス
正式名称はカエノラブディティス・エレガンス。非寄生性の線虫で、土壌中に生息する。分子遺伝学的解析に使用されており、全神経細胞の接続地図や遺伝子発現プロファイルが公表されている。ヒトの遺伝子数と同等に約2万個の遺伝子を持ち、類似の遺伝子も多い。
注2)
LHT1遺伝子
シロイヌナズナの高温耐性の多様性に関わる遺伝子Long-term Heat Tolerance1 (LHT1) /MAC7。LHT1/MAC7遺伝子は、mRNAのスプライシングに関与するRNAヘリカーゼをコードし、LHT1/MAC7はヒトのAquarius (AQR)の相同遺伝子である。
注3)AQR(Aquarius)
ヒトや他の真核生物に存在するタンパク質で、特にスプライソソームの機能に関与する。
注4)イントロン
遺伝子のDNAやその転写産物であるRNAの中で、エクソンに挟まれた部分であり、最終的な成熟mRNAには含まれない非コード領域。
注5)スプライシング、スプライシング反応
真核生物の遺伝子発現過程において、mRNA前駆体からイントロンを除去し、エクソンを連結して成熟mRNAを生成する過程と反応。この過程は、遺伝子から正確で機能的なタンパク質を生成するために不可欠である。
注6) RNAシーケンシング解析
細胞内のRNAの量と種類を高精度に測定する技術。細胞からRNAを抽出し、cDNAに変換し、次世代DNAシーケンサー(NGS)を用いてcDNAの塩基配列を決定し、遺伝子発現のプロファイルを作成することで、遺伝子発現の変動などを測定することができる。
注7)酸性スフィンゴミエリナーゼ
細胞膜やリソソームなどに存在する酵素で、スフィンゴ脂質のスフィンゴミエリンを加水分解してセラミドとリン酸化コリンを生成する。セラミドを含むスフィンゴ脂質は、細胞膜のシグナル伝達の中心となる脂質ラフト(lipid raft)の形成にも関与する。
注8)リン脂質スクランブラーゼ
細胞膜やミトコンドリアなどの脂質二重膜に存在し、リン脂質(ホスファチジルセリン)を内膜から外膜へと移動させる。SCRMの主な活性化因子はCa2+であり、これは血液凝固、アポトーシス、マイトファジーシグナルによって誘導される。
注9)ヘリカーゼ、ヘリカーゼドメイン
ヘリカーゼは、DNAやRNAの二重らせん構造を解きほぐすタンパク質。らせん状のDNAやRNAを分解し、一本鎖の状態に戻す酵素。ヘリカーゼ活性をもつタンパク質のドメイン構造がヘリカーゼドメイン。ヘリカーゼは他の核酸関連酵素と共同して働く。
注10)スプライソソーム
真核生物の細胞内で、mRNA前駆体(pre-mRNA)のイントロンを除去し、エクソンを連結するスプライシング反応を行う大きな複合体を指す。この複合体を構成する分子がスプライソソーム因子である。スプライソソームの働きにより、成熟したmRNAが生成され、mRNAが翻訳されて機能的なタンパク質が作られる。
<掲載論文名>
“The intron binding protein EMB-4 is an opposite regulator of cold and high temperature tolerance in
Caenorhabditis elegans”
(イントロン結合タンパク質EMB-4は線虫の低温および高温耐性の相反的な制御因子である)
英国科学雑誌「
PNAS nexus」Volume 3, Issue 8, page293
DOI: doi.org/10.1093/pnasnexus/pgae293
https://doi.org/10.1093/pnasnexus/pgae293
<発表者> *責任著者、#共同筆頭著者
*#太田 茜(オオタ アカネ) (甲南大学理工学部 特任研究准教授)
#佐藤 夕希(サトウ ユキ)(甲南大学大学院自然科学研究科 大学院生(研究当時))
礒野一帆(イソノ カズホ)(東京農業大学大学院生命科学研究科 大学院生(研究当時))
梶野拓磨(カジノ タクマ)(東京農業大学大学院生命科学研究科 大学院生)
田中啓介(タナカ ケイスケ)(東京農業大学 生物資源ゲノム解析センター 助教)
太治 輝昭(タジ テルアキ)(東京農業大学大学院生命科学研究科 教授)
*久原 篤(クハラ アツシ)(甲南大学理工学部/大学院自然科学研究科/統合ニューロバイオロジー研究所、教授)