横浜市立大学大学院医学研究科免疫学 山﨑貴弥さん(博士課程4年)、西山晃准教授、田村智彦教授らの研究グループは、熊本大学、国立遺伝学研究所、米国国立衛生研究所と共同で、病原体やがんに対する免疫に関わる樹状細胞
*1の分化に必要な転写因子
*2 IRF8の発現を制御するエンハンサー
*3群の相互作用メカニズムを明らかにしました。
本研究成果は、米国科学雑誌「Cell Reports」に掲載されました。(2024年4月12日オンライン先行公開)
研究成果のポイント
・病原体やがんに対する免疫に関わる転写因子IRF8をコードする遺伝子の発現を制御する複数のIrf8エン
ハンサー間の相互作用を解析した。
・各エンハンサーが、分化段階によって異なるメカニズムで他のエンハンサーを活性化していることが分
かった(図1)。
・この発見は細胞分化
*4における普遍的な原理に関わっている可能性がある。
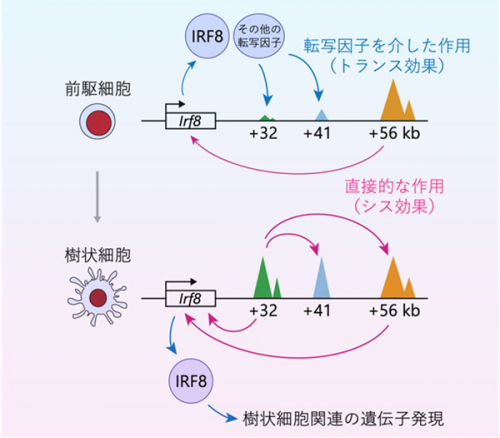
図1 樹状細胞分化におけるIrf8エンハンサー相互作用の概念図
研究背景
ウイルスや細菌などの病原菌から身体を守る免疫細胞である白血球は、造血幹細胞
*5から多能性造血前駆細胞
*6などの前駆細胞段階を経て、特定の成熟細胞へと分化します。樹状細胞や単球を含む単核貪食細胞
*7の分化では転写因子IRF8が重要な役割を果たします。本研究グループでは前駆細胞で転写因子IRF8が高発現すると樹状細胞、低発現では単球、発現しないと好中球へと分化することを報告してきました。樹状細胞への分化におけるIrf8発現量は3つのエンハンサー(+56 kb、+41 kbおよび+32 kbエンハンサー)が順番に機能することよって制御されていることも見出しています(Murakami et al, Nat Immunol, 2021など)。しかし、これらIrf8遺伝子のエンハンサー群の物理的・機能的な相互作用は不明でした。一般に遺伝子は複数のエンハンサーによってその発現が制御されることが多いのですが、エンハンサー間の相互作用には不明な点が多く、細胞分化のメカニズムにおいても根元的な問いの一つとなっています。
研究内容
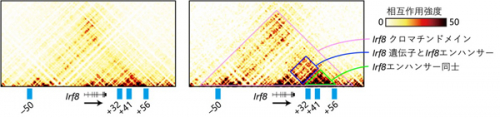
まず初めに、樹状細胞分化でのクロマチン高次構造
*8の変化を解析しました。Irf8遺伝子を含む大きなクロマチンドメイン
*9が多能性造血前駆細胞から形成され、樹状細胞の分化に伴い、Irf8エンハンサー同士やIrf8遺伝子領域との結合が増強することがわかりました(図2)。
図2 Irf8遺伝子領域におけるクロマチンの物理的相互作用をTiled-Cという手法で測定した。大きなIrf8クロマチンドメインは多能性前駆細胞でも見られるが、Irf8遺伝子とIrf8エンハンサーおよびIrf8エンハンサー同士の相互作用は樹状細胞において増強されている。
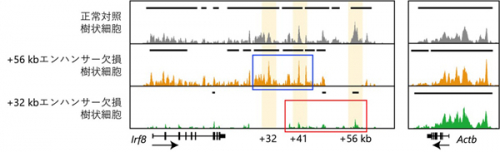
さらに、Irf8エンハンサーの機能的な相互作用の解明を試みました。2つあるIrf8遺伝子対立遺伝子座
*10のうち両方のIrf8エンハンサーを欠損させると樹状細胞が産生されず解析できないため、一方のIrf8エンハンサーのみを欠損させたヘテロ欠損マウスを用いました。その結果、Irf8エンハンサーがお互いに活性化し合っている可能性が浮かび上がりました。この相互活性化は、同一の対立遺伝子座における物理的な結合による直接的な作用(シス効果)に加え、IRF8発現誘導を介してもう一方の対立遺伝子座にも働く間接的な作用(トランス効果)によって生じている可能性が考えられます。それらを区別するために、両方の遺伝子座のIrf8エンハンサーを欠損した前駆細胞にIRF8を人為的に発現させ樹状細胞への分化を救済して、他のエンハンサーの状態を解析しました。その結果、多能性前駆細胞段階から機能する+56 kbエンハンサーはIRF8の発現を介した間接的なトランス効果によって下流のIrf8エンハンサーを活性化するのに対して、樹状細胞への分化が決まった後に機能する+32 kbエンハンサーは直接的なシス効果によって上流のIrf8エンハンサーの活性化を維持していることが明らかになりました(図3)。
図3 Irf8エンハンサー欠損前駆細胞にIRF8を人為的に発現させ分化を救済した樹状細胞をCUT&Tagという手法でヒストンのアセチル化を測定して、残存Irf8エンハンサーの活性を評価した。+56 kbエンハンサーを欠損した樹状細胞では残存Irf8エンハンサーが活性化状態にある(青い四角)のに対して、+32 kbエンハンサーを欠損した樹状細胞では残存Irf8エンハンサー(赤い四角)は活性化していない。
今後の展開
本研究では、Irf8遺伝子の発現制御を誘導する複数のエンハンサー間の相互活性化が、分化段階によって異なるメカニズムで生じていることが分かりました(図1)。これにはどのような意味があるのでしょうか。直接的なシス効果は物理的に結合すれば確実に生じるのに対し、間接的なトランス効果は複数の転写因子のネットワークによって生じるためより柔軟に調節される余地があります。早期の多能性前駆細胞段階から機能する+56 kbエンハンサーがもし直接的なシス効果で下流の+41 kbや+32 kbのエンハンサーを自動的に活性化してしまうと、前駆細胞は全てIRF8を高発現して樹状細胞に分化してしまい、IRF8の発現が低いあるいは無い時に産生される単球や好中球には分化できなくなってしまうでしょう。すなわち、複数の細胞種への分化能を持つ前駆細胞と、運命がすでに決定された細胞とでは、エンハンサー相互作用の様式が異なることはとても合目的と考えられます。このような調節は他の細胞種への分化においても用いられている普遍的な仕組みであると予想され、更なる研究でそれが明らかになることが期待されます。
研究費
本研究は、文部科学省、日本学術振興会、日本血液学会による研究助成と、本学先端医科学研究センターにおける文部科学省特色ある共同利用・共同研究拠点「マルチオミックスによる遺伝子発現制御の先端的医学共同研究拠点」ならびに文部科学省先進ゲノム支援(PAGS)の支援を受けて実施されました。
論文情報
タイトル: Physical and functional interaction among Irf8 enhancers during dendritic cell differentiation
著者: Takaya Yamasaki*, Akira Nishiyama*, Nagomi Kurogi, Koutarou Nishimura, Shion Nishida, Daisuke Kurotaki, Tatsuma Ban, Jordan A. Ramilowski, Keiko Ozato, Atsushi Toyoda, Tomohiko Tamura (*Co-1st authors)
掲載雑誌: Cell Reports
DOI:
https://doi.org/10.1016/j.celrep.2024.114107
用語説明
*1 樹状細胞:免疫応答の初期段階でサイトカインなどを分泌する自然免疫を生じるのに加え、T細胞に異物の目印(抗原)を提示することによって獲得免疫の引き金をも引く、免疫の司令塔と表現される細胞。
*2 転写因子:それぞれ特徴的なDNA配列を認識して様々なゲノム部位に結合することで、遺伝子からRNAの転写を制御するタンパク質。ヒトでは1,500種類以上あると言われている。
*3 エンハンサー:転写因子が結合することで活性化が制御され遺伝子の発現を調節するゲノムDNA領域。
*4 細胞分化:様々な種類の細胞に変化できる細胞(幹細胞や前駆細胞)が、特定の機能を持って成熟した細胞になっていくこと。
*5 造血幹細胞:骨髄内に存在し、全ての血液細胞に分化する能力と、細胞分裂しても自らを維持する自己複製能を有する組織幹細胞の一種。
*6 多能性造血前駆細胞:自己複製能は低いが多種類の血球細胞を作り出す能力を持つ前駆細胞。
*7 単核貪食細胞:単球・マクロファージと樹状細胞の総称。これらの細胞は貪食能や炎症性サイトカイン産生、抗原提示能などの機能を有しており、自然免疫のみならず獲得免疫に繋ぐ重要な免疫細胞群である。
*8 クロマチン高次構造:遺伝子発現に重要とされるクロマチン(ゲノムDNAとヒストンなどのタンパク質の複合体)の立体構造。
*9 クロマチンドメイン:クロマチン高次構造において、会合頻度の高いクロマチンの構造単位。
*10 対立遺伝子座:染色体上の遺伝子の位置を、遺伝子座という。ヒトをはじめとした2倍体の生物は、父親と母親それぞれから引き継いだ同一の遺伝子があり、2倍体の生物は各遺伝子について2つの対立遺伝子座を持つ。
参考文献
Murakami et al; A RUNX–CBFb-driven enhancer directs the Irf8 dose-dependent lineage choice between DCs and monocytes. Nat Immunol 22(3): 301-311(2021)